5 Gynogenetic Fish
evolution, coevolution, host-parasite interactions, Muller’s ratchet, parthenogenesis, Red Queen hypothesis, sexual reproduction
New Zealand felt like home after five years, and we had many good friends and colleagues in Christchurch, especially David Lloyd. David and I talked often about strategic models of natural selection (game theory), of which he was one of the world’s leaders. I was particularly influenced by his idea that sexual reproduction, polymorphism, phenotypic plasticity, and hermaphroditism could all be seen under the umbrella of variation strategies
(Lloyd 1984). Sadly, when Lynda completed her PhD, her visa expired, and we had to leave New Zealand. But where to go?
Lynda was a rising star when she completed her PhD. She landed a post-doc at Rutgers University to work with Tom Meagher, so we moved to New Jersey in January 1989. Once again, Lynda was supporting us. Steven Handel gave me a desk and some space in his lab. Soon afterward, Peter Morin asked me to give a seminar on sex/asex at Rutgers. I knew of Peter’s pioneering work in community ecology for which he had received a young investigator’s award from the Ecological Society of America (the Mercer Award). I was quite nervous to give a seminar in front of one of my heroes. Another of my heroes, Professor Bob Vrijenhoek, was also in the audience. As described in Section 2.2, Bob was the creative mind behind the Frozen Niche-Variation Hypothesis, and he was well known for his work on sexual and asexual fish (topminnows in the genus Poeciliopsis). He was also the director of the Center for Theoretical and Applied Genetics (CTAG) at the New Brunswick campus of Rutgers University, and he was the Editor-in-Chief of the journal Evolution, the most prestigious journal in evolutionary biology. After my talk, Bob, Peter, Lynda, and I went out for dinner. Somewhere in the middle of the meal, Bob offered me a post-doc to work with him at CTAG. I was dumbfounded. But I managed to say yes, When do I start?
He said, How about tomorrow?
For this to make sense, you need to know more about Vrijenhoek’s fish. Like the snails, the fish have coexisting sexual (diploid) and asexual (triploid) forms. The asexual fish, however, were gynogenetic,
which means that the eggs require fertilization to kick-start embryogenesis. The sneaky eggs, however, do not generally incorporate the sperm’s DNA (review in Vrijenhoek 1998).1 The outcome is the coexistence of sexual and asexual lineages within the same semi-isolated populations.
Bob had worked on these fish for decades, focusing on small streams in Mexico. He had worked in particular on the coexistence of a sexual form of the fish, Poeciliopsis monacha, with two gynogenetic clones, which were interspecific hybrids between P. monacha and P. lucida (Vrijenhoek 1978, 1979). The triploid clones were independently derived from their sexual ancestors, with two copies of the monacha genome and one copy of the lucida genome (Schultz 1969). During his studies, Bob had frozen fish for many years from several of his sites. He remembered that some of the fish in his freezer were heavily infected with black-spot disease. The black spots are clearly visible on the body of the fish. The spot itself is caused by trematode larvae that burrow into the fish and encyst (genus Uvulifer). The fish’s immune system then coats the cyst with melanin, which turns the cysts black. Bob thought it would be interesting to see if the asexual fish were more infected than the sexuals. The prediction under the Red Queen Hypothesis was that the most common asexual clones would be most infected by black spot disease (Section 5.1).
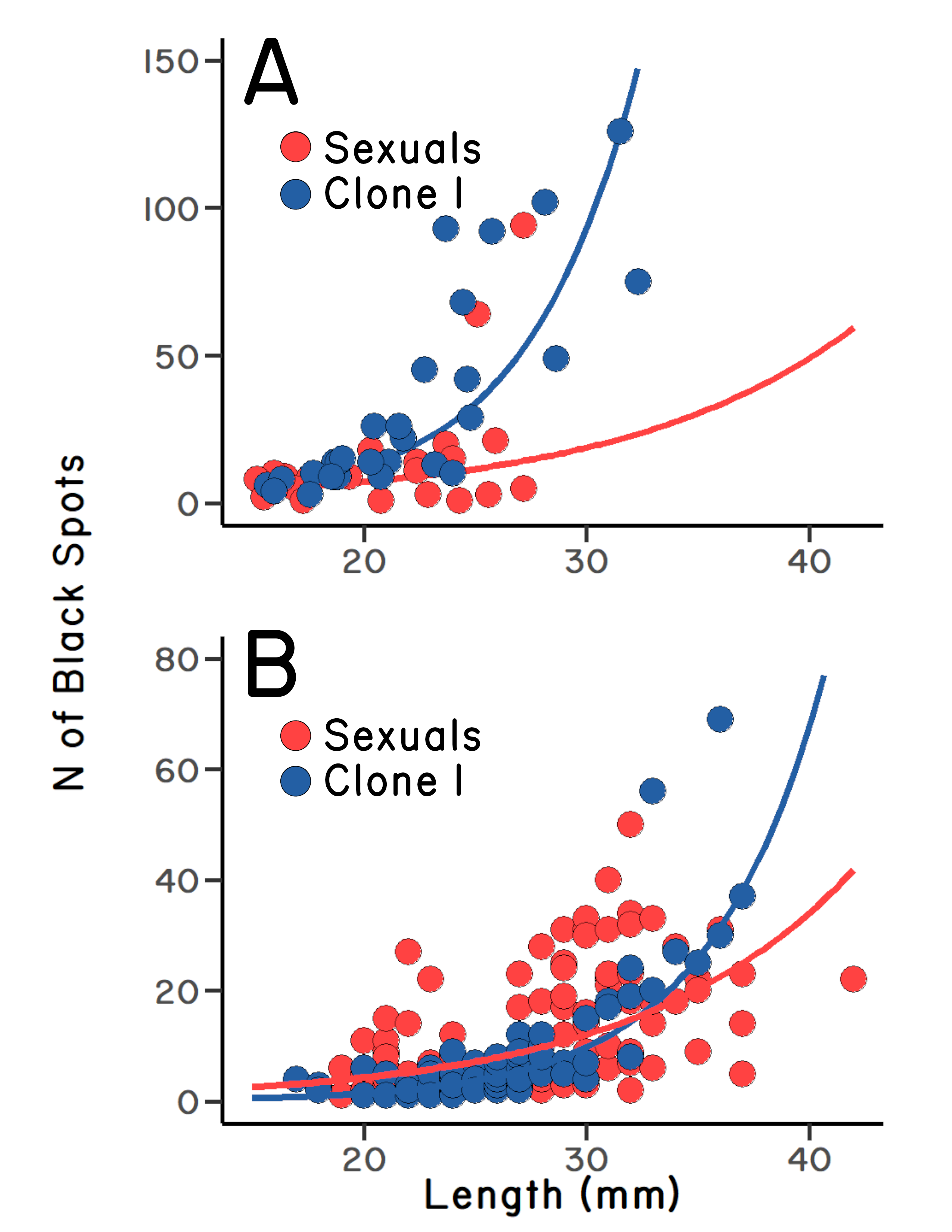
Bob enlisted the help of one of his PhD students, Clark Craddock. Clark ran the allozyme electrophoresis, which was necessary at the time to tell sexual fish from the two clones. She also helped count the cysts, run the statistics, and write the paper. We found right away that larger fish had more cysts, which makes sense. But we also found that the most common clone, clone 1, had significantly more black spots per unit body length than the sexual fish in Log Pool (Figure 5.1 A & B). In addition, the sexual fish showed greater size-corrected variation in the number of parasitic cysts than the asexual fish, which makes sense if the sexual population contained more genetic variation for resistance to infection. So, after correcting for size, the sexual fish were both less infected and showed more variation in infection than the clone 2.2 This seemed very promising for the Red Queen.
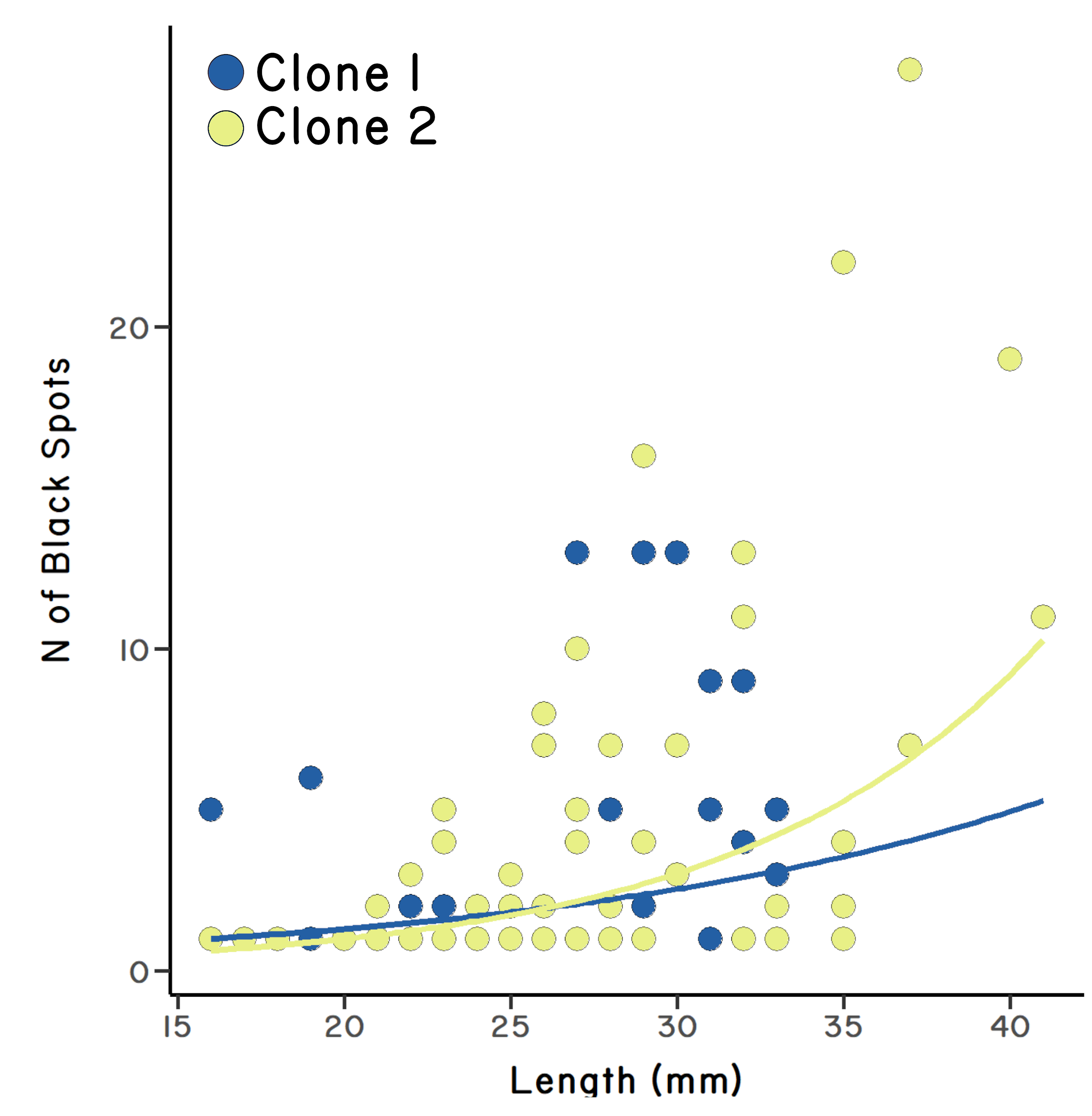
Similar results were observed in a second pool (Sandal Pool) in which both clones coexisted. In this pool, clone 2 was more common than clone 1, and it was also more infected than clone 1 (Figure 5.2). However, clone 1 was not more infected than the sexual form of P. monacha, which showed that clone 1 is not inherently more susceptible to infection (Lively et al. 1990). Taken together, the results suggest that the parasites had evolved to disproportionately attack the most common local clone.
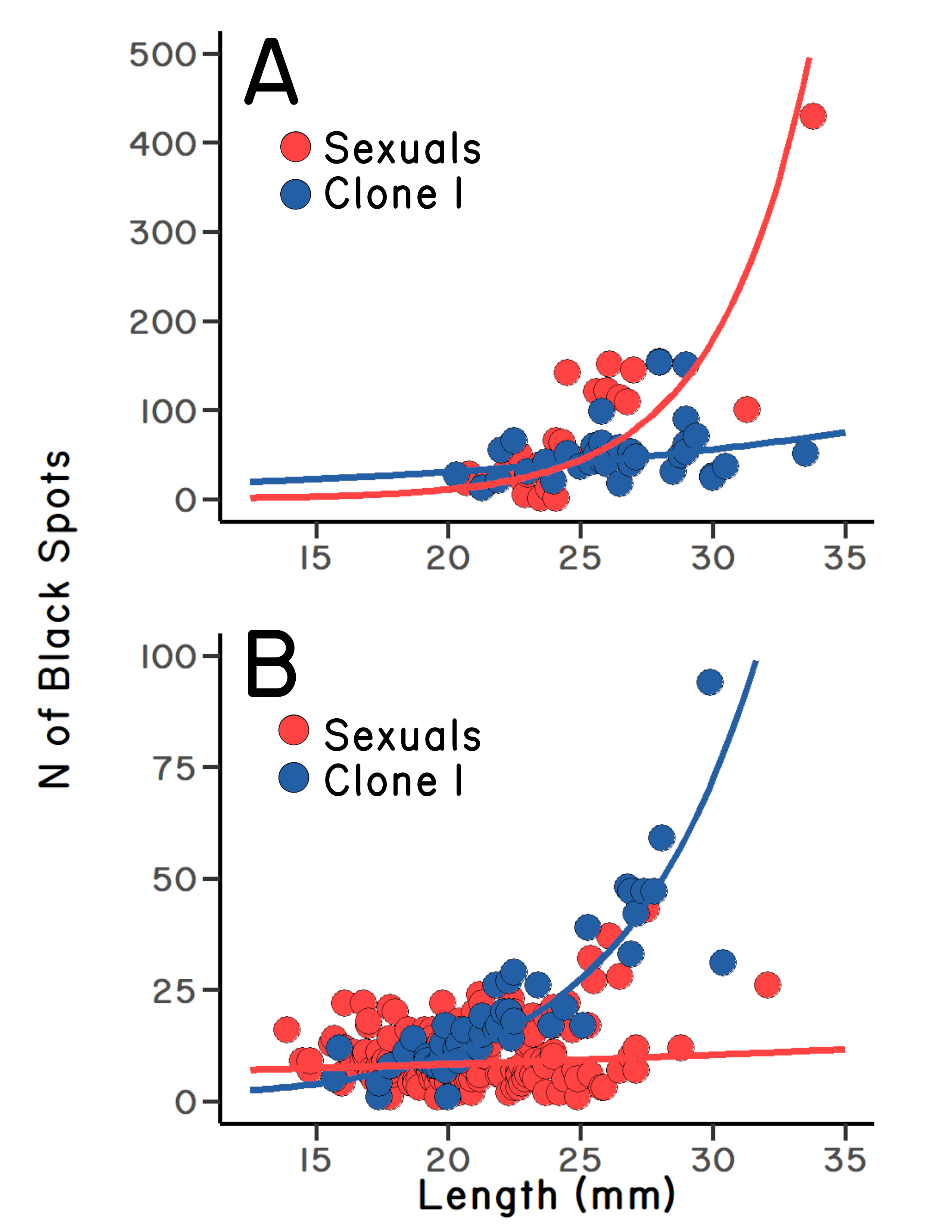
But we had more samples to run. When we examined another pool, the trend reversed: sexual fish were more infected than asexual fish (Figure 5.3 A). We were confused. I remember Clark telling Bob of this new result. He said something like, Well, that’s science,
and then headed back to his office. He was halfway there when he turned around and asked, what sample was that?
Clark told him, Heart Pool, 1983.
Bob got a big smile. Then he told us that, in that sample, the sexual fish were highly inbred. The population had descended from a small number of founders, and they were homozygous for most loci. So, the sexual fish were more infected in that sample, but they were also genetically depauperate. Under the Red Queen, there is no value to sexual reproduction unless there is genetic variation in the population. On the other hand, the greater level of infection in sexual fish might have been caused by inbreeding depression, which was known for other fitness-related traits (Vrijenhoek & Lerman 1982).
It was an amazing set of results, but there was another twist. Bob had added some fish (in 1983) to the isolated founder population in Heart Pool. His goal was to increase the genetic diversity of the sexual population. After a few years, he resampled the site, and fortunately, he had saved the samples in his freezer from 1985. Clark and I had a look. The pattern had reversed: now the sexual fish were less infected than the asexual fish (Figure 5.3 B). In addition, the size-corrected variance in infection had increased in the sexual fish in just two years. The advantage to outcrossing seemed to depend on genetic diversity.
In summary, the data suggested that the parasites had evolved to infect the most common local host clone in the different pools. They also suggested that sex in genetically diverse host populations results in protection from disease. Both results were consistent with the Red Queen.3 Proof? No. Fascinating? Yes. In any case, looking back, I was very lucky to have had the opportunity to work with Professor Vrijenhoek.
5.1 Appendix: Within Versus Between Populations
It can be a bit confusing, but I think that the predictions of the Red Queen Hypothesis depend on whether one is looking within versus between populations. In Chapter 3, I suggested that snail populations having more parasites should be more likely to have some sexual reproduction. Here I am suggesting that asexual fish would be expected to have more parasites. How does that work?
Between populations we would expect to find that host populations without coevolving parasites would evolve to reproduce by parthenogenesis. As such, sex should be positively correlated with infection prevalence. The pattern, however, would be expected to be messy, even in cases where parasites are the major driving force for sex (Lively et al. 2021). Nonetheless, the common expectation is that sex should be more common in host populations having a history of strong parasite-mediated selection against common genotypes.
Within populations the most common host genotypes should be more infected, at least periodically. Hence the most common clonal genotypes should be more infected than cross-fertilizing hosts in the sexual population. This might be especially true in cases such as Vrijenhoek’s fish discussed here, where parasites may not be virulent enough to drive strong oscillatory dynamics in clone frequencies, and sexual reproduction is most likely maintained by sperm-dependent (pseudogamous) reproduction in the clones (Moore & McKay 1971) and/or by greater niche width in the sexual population (Schenck & Vrijenhoek 1986).
Paternal leakage of DNA in otherwise asexual vertebrate species now seems possible (review in Lampert & Schartl 2010).↩︎
In a follow-up study (using different samples), Steve Weeks (1996) did not find some of our significant effects. His sample sizes, however, were small, suggesting that his power to detect differences was low.↩︎
It is important to point out that we were not claiming that parasite-mediated selection is maintaining sex in this system. For example, we wrote,
[W]e do not intend to imply from this analysis that the trematode is selecting for the maintenance of sex in these fish.
Niche partitioning and/or the sperm-dependence of the gynogenetic females would most likely ensure the persistence of sexual individuals (Moore & McKay 1971; Schenck & Vrijenhoek 1986). Our claim was simply that parasites had evolved to disproportionately infect the most common clone unless the sexual population was highly inbred.↩︎